

Effect of brown rice fermentation
The nine brown rice varieties were evaluated for P. acetolactic MNL5. FBR-1741 exhibited the highest pancreatic lipase inhibitory activity among the nine brown rice varieties at the 48 h optimized incubation time. Furthermore, the strongest TPC, TFC, and antioxidant activities were also observed. In earlier investigations, fermenting diverse substrates increased the bioactive chemicals and anti-obesity potential in the MNL5 strain. Fermentation stimulates the biochemical processes of bacteria, which break down the cell wall structurally, producing various bioactive metabolites16.
Impact of FBR on lipase inhibition
The inhibitory activity of the pancreatic lipase enzyme and the associated compounds is shown in Fig. 1a, the fermentation process enhanced the lipase activity of brown rice compared to that of other rice varieties. Lipase inhibitory activity was highest in 1741 (87.6 ± 2.1%) > 1708 (79.2 ± 1.6%)>, while DM25 (62.5 ± 1.9%) and DM29 (59.5 ± 2.1%) showed enhancement. Lipase inhibition is an efficient technique for preventing the absorption of triacyl glycerides in people who suffer from hypercholesterolemia. Fermented materials suppress pancreatic lipase, although few reports have been published. Moreno et al.20 observed 80% lipase inhibition with 1 mg/mL grape seed extract. In addition, Wang et al.21 revealed that NTU 101-fermented green tea promotes lipase activity in adipocytes to limit lipid droplet formation and increase lipolysis.
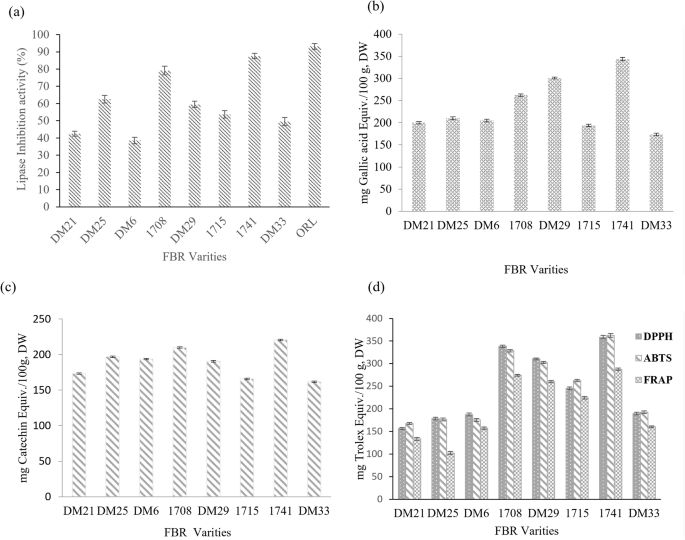
a Anti-obesity in vitro pancreatic lipase inhibitory activity of fermented Brown rice extracts. b Total phenolic content (TPC). c Total flavonoid content (TFC). d Antioxidant activity. Each bar represents the mean ± SD of triplicate values. https://doi.org/10.6084/m9.Figureshare.24781476.
Total phenolic and flavonoid content of FBR
Figure 1b, c shows the TPC and TFC of nine varieties of fermented brown rice. TPC and TFC increased in FBR-1741 compared to the other FBR varieties. After comparison, FBR-1741 had the highest total phenolic and flavonoid content (TPC-343.5 ± 3.89 mg GAE equiv./100 g, DW; TFC-220.5 ± 3.81 mg GAE equiv./100 g). The second improved TPC and TFC content was FBR 1708 (Fig. 1b, c). The overall phenolic and flavonoid content is anticipated to benefit the antioxidant activity. Our findings suggest that more work is needed to boost the bioactivity of polyphenolics in FBRs and achieve antioxidant efficiency. Most cereals contain esterified connections to the matrix of the grain wall due to a lack of phenolic compounds21. Fermentation is a possible method for increasing the bioavailability of grain phenolics22. The above method may release insoluble or bonded phenolic substances. The current study compared nine varieties with fermentation using MNL5 and found that FBR-1741 enhanced polyphenols compared to other brown rice varieties from Korean resources.
The effect of DPPH, ABTS, and FRAP scavenging activity
The enhanced antioxidant properties of brown rice extracts have been attributed to reducing capacity, metal ion chelation, and radical scavenging, among other mechanisms23. Absorption spectroscopy is a popular method for measuring the antioxidant activity of natural materials. FBR-1741 (358.5 ± 2.8 mg Trolox equiv./100 g, DW) and FBR-1708 (338.5 ± 3.1 mg Trolox equiv./100 g, DW) had the highest DPPH values. With the lowest results (156.5 ± 2.5 mg Trolox equiv./100 g, DW), FBR-DM21 was tested. Therefore, ABTS is necessary to assess the radical scavenging ability of grain. While the FRAP test was primarily designed to measure antioxidant capacity in plasma, it is now routinely used to analyze antioxidant capacity in isolated chemicals and biological materials24. It recognizes changes in absorbance brought on by blue iron (II) derived from iron oxide (III). The highest ABTS activity was observed in FBR-1741 (362.5 ± 3.3 mg Trolox equiv./100 g, DW). In addition, FBR-DM21 has the lowest ABTS activity. Similar results with DPPH were obtained in this study’s ABTS and FRAP tests. MNL5 FBR-1741 contained the maximum levels of FRAP (295.53.3 mg Trolox equiv./100 g, DW), followed by 1708 and DM21 (Fig. 1d). MNL5 FBR-1741 had the highest activity level out of the nine samples, according to the findings of the antioxidant assays. These results were greater than those reported previously by IIowefah et al.25 for fermented brown rice flour. The bioavailability of phenolics and flavonoids, as well as antioxidant activity, can be measured using this method, revealing variations in the extracts’ antioxidant activity. In addition, their chemical composition strongly influences the quenching power of the phenolic compounds. Antioxidants help prevent aging, obesity, and diabetes, which have been shown to be associated with the action of free radicals, as well as the degradation of essential fatty acids26. In recent years, fermentation has been regarded as an efficient method for enhancing the antioxidant activity of grains. Landete et al.27 reported the glycosidase activity of L. plantarum in relation to enhancing the bioaccessibility and bioavailability of dietary phenolic compounds and resulting in greater antioxidant activity. Our results show that FBR-1741, which was evaluated by DPPH, ABTS, and FRAP methods, has the strongest antioxidant activity.
UHPLC-Q-TOF-MS/MS to identify the polyphenols in fermented brown rice
Our study used METLIN and the Metabolomics Workbench to assist us in determining the classification of polyphenolics. Table 1 shows that 17 phenolic substances were found in the ethanolic extracts of FBR-1741. This study obtained high amounts of phenolic substances formed in optimized conditions within 48 h. In addition, after 48 h of fermentation, ferulic acid, cinnamic acid, p-coumaric acid, protocatechuic acid, ethyl 4-hydroxybenzoate, caffeic acid, homovanillic acid, butylparaben, gallic acid, quercetin, isorhamnetin, sophoricoside, phenprobamate, daphnetin, cantharidin, genipin, and irisflorentin were identified. UHPLC-Q-TOF-MS/MS analysis identified 17 components in FBR, including 11 phonic compounds, four flavonoids, and two organic acids (Supplementary Fig. 1a–d). All of the top 17 active metabolites, especially ferulic acid, quercetin, isorhamnetin, irisflorentin, and protocatechuic acid, have a role in maintaining lipid metabolic stability. However, the FBR-1741 type exhibited the greatest level of phenolic compounds found. Based on our study, UHPLC-Q-TOF-MS/MS reveals the enhanced polyphenolic metabolites for FBR-1741 for anti-obesity functionality. In addition, 1708 shows 14 phenolic substances were found in the ethanolic extracts (Supplementary Table 1). Kuppusamy et al.28 observed that FA was imperative for antiadipogenic and lipogenic effects by regulating key adipocyte factors and enzymes and accelerating lipolysis via the HSL/perilipin mechanism. In addition, another investigation showed that ferulic acid reduced the risk of NAFLD and was developed as a functional food or beneficial substance28. In our prior study, fermented onion lowered cholesterol and enhanced quercetin to improve pancreatic lipase activity for synergistic anti-obesity benefits16. Isorhamnetin suppressed body fat and enhanced fat oxidation in the NHR-49-dependent pathway by lowering fat accumulation in a C. elegans study29. Jung et al.30 found that caffeic acid may reduce hyperglycemia by stimulating insulin secretion and reducing insulin resistance in db/db mice. Quercetin also improved the insulin resistance index, antioxidant enzyme activity, and insulin signaling in high-fat-diet-fed animals, showing its free-radical scavenging properties31.
Network pharmacology relations for FBR-1741 metabolites
In the present study, 132 common target genes for obesity were acquired from available databases such as TCMSP, DisGeNET, and GeneCards (Supplementary Fig. 1a). The obesity-associated target genes interaction with 17 active metabolites chosen from FBR is provided in Supplementary Table 2. Drug screening criteria parameters, including OB, DL, and BBB, were selected for these 17 active compounds. The obesity-related target gene (132) interaction network was constructed using the STRING database with a high confidence score (0.700) (Supplementary Fig. 1b). The major hub proteins are vital in controlling and expressing all interacting sub-proteins to carry out their biological and molecular functions at any time. Five top-ranked hub proteins (VEGFA, AKT1, JUN, IL-6, and MMP9) were associated with the target disease–gene interaction network (Supplementary Fig. 1c). These selected hub proteins may be controlled/regulated by the biological and molecular functions of the remaining disease-associated target genes. According to the DisGeNET and GeneCards database analyses, five compounds showed the best drug-likeness networking properties against the five hub proteins. The bioactive FBR compounds (17) interacted with five hub proteins and the remaining disease-associated target genes. Among the 17 FBR compounds, ferulic acid, quercetin, isorhamnetin, protocatechuic acid, and irisflorentin had significant interactions with the top five hub proteins (Fig. 2). These results indicated that these FBR compounds might regulate or influence the genes related to obesity. Authenticating the hypothesis that VEGF-A concentration in serum is strongly associated with body mass index, elevated levels of VEGF-A were detected in humans as well as animal models that have become overweight or obese32. Adipose-VEGF stimulation reverses a cascade of events—adipocyte death, hypoxia, inflammatory processes, abnormal fat accumulation, and lipotoxicity—in chronically confronted animals and maintains glucose balance. According to Robciuc et al.33, inhibiting VEGFR-2 (KDR gene-encoded protein) activation might lower obesity by inhibiting angiogenesis and reducing fat mass. Shearin et al.34 showed that AKT1 signaling downstream of insulin and IGF-1 receptors regulates fat formation and regulation. IL-6 is an inflammatory factor that may predict insulin resistance and cardiovascular disease and contribute to their development. It is connected to abdominal adipose tissue and may influence TNF-α and other inflammatory factors. IL-6 and TNF-α are known to be produced by adipose tissue and may contribute to body mass augmentation35. According to Wang et al.36, network pharmacology-guided baicalin may reduce obesity by upregulating SLC2A1 and downregulating TNF, NFKB1, SREBF1, PPRGA, and CASP3. The current work focuses on the strong connections that VEGFA, AKT1, JUN, IL-6, and MMP9 hub genes for obesity-related functions have with the FBR-1741 metabolites.

The blue table nodes represent the 17 components, the red diamond represents the hub-gene, and the green circle nodes represent the corresponding 132 target genes of the ingredients. https://doi.org/10.6084/m9.Figureshare.24781479.
Performance of molecular docking
The five active FBR compounds were subjected to molecular docking with the top hub proteins. These compounds demonstrate the highest interaction with hub proteins. The binding scores, amino acid residues, and hydrogen bond distances are represented in Supplementary Table 3. Quercetin demonstrated the highest binding scores against the AKT1 (−6.8), IL6 (−7.2), and MMP9 (−8.0) hub proteins. Three amino acid residues, A: ARG169, A: MET68, and A: GLU173, with hydrogen bond distances of 2.544, 2.304, and 2.758 Å, respectively, were found against IL6. Isorhamnetin shows better interaction scores against the VEGFA (−8.5) and JUN (−5.3) hub genes than the other FBR compounds. Four hydrogen bond interactions with a strong h-bond distance were found against VEGFA (C: TYR357–2.243 Å; C: HIS415–2.059 Å; B: TYR299–2.918 Å and B: ILE418–2.406 Å). Irisflorentin had a better docking score against VEGFA (−7.4) and three amino acid residues with a strong hydrogen bond distance (Supplementary Table 3). Furthermore, ferulic acid demonstrated moderate docking scores against all the hub proteins. The two best hydrogen interactions were found with each hub protein except the IL6 gene. Among these, VEGFA showed a better binding score of −6.9 with the two hydrogen bond interactions and a better bond length (B: ASP323–2.315 Å and B: ILE418–2.970 Å) (Supplementary Fig. 3, Supplementary Table 2). Protocatechuic acid docked with MMP9 showed a better binding score of −7.1 and five different amino acid residues with hydrogen bond distances (Table 2). The docking results demonstrated that quercetin, isorhamnetin, Irisflorentin, ferulic acid, and protocatechuic acid are potential regulators of obesity-associated hub genes. The mechanisms through which ferulic acid inhibits α-amylase and α-glucosidase were investigated by Li et al.37. These five compounds revealed the highest docking scores and hydrogen bonding interactions out of 17 FBR bioactive metabolites.
Quantification of network pharmacology and molecular docking-guided metabolites
In silico-determined metabolites were detected in the RBR and FBR (1708 and 1741) samples to assess the efficacy of fermentation on polyphenolic content (Fig. 3). Since ferulic acid has been shown to improve metabolic health in several ways, it was considered in this investigation. After 48 h of FBR-1741, ferulic acid was significantly higher than in the initial sample and other varieties (RBR-1741: 175.2 µg/g; FBR-1741: 570.2 µg/g). Doses for future investigations based on quantification and metabolites.

Heat map represent the levels of ultra high-performance liquid chromatography (UHPLC) polyphenolic metabolites in raw brown rice compared to fermented brown rice (FBR) varieties, FBR-1708 and FBR-1741; raw brown rice (BR) BR-1708 and BR-1741 at a concentration of 1 mg/mL.
In vivo C. elegans correlation of lifespan and lipid reduction mechanism by FBR extracts with metabolites
High-glucose diets may shorten the lifespan of C. elegans. The mean and maximum longevity (34.5 ± 1.5%) of the FBR-1741 groups were significantly longer than those of the other brown rice groups and those of the negative control groups (30.2 ± 1%). Based on network analysis, we evaluated the lifespan for the FBR-guided top five metabolites from network pharmacology, such as ferulic acid (31.5 ± 1.4), quercetin (28.5 ± 1), isorhamnetin (11.5 ± 09), irisflorentin (22.5 ± 0.8), and protocatechuic acid (6.5 ± 1). The lifespan was prolonged in the ferulic acid-supplemented diet group compared to the other metabolite groups. Ferulic acid, directly associated with a longer lifespan, was also elevated by FBR1741 (Fig. 4a). Obesity-induced N2 C. elegans consumed OP50 and glucose (PC) groups and died on the 13th day. Our results confirmed those of a previous investigation that suggested that FBR (mixed variety) might increase longevity following a high glucose diet16. Many fermented materials demonstrate that lipid metabolism regulates C. elegans longevity by connecting apoptosis, embryonic stem cells, and chromosomal factors. According to Hou et al.38, zymolytic grain extract increased longevity dose-dependently compared to the controls. This research shows that extracts from MNL5 FBR-1741 protect against the shorter lifespan caused by hyperglycemia by reducing fat accumulation.

a Lifespan analysis. b C. elegans lipid droplets visualized by Nile-red staining and Oil Red. c mean of fluorescence intensity measured by Image J software; NC-OP50, PC-OP50 + Glucose, DC-OP50 + Glucose + Orlistat, FBR1741-OP50 + Glucose + FBR1741 (1 mg/mL), FA-OP50 + Glucose + FA, QU-OP50 + Glucose + QU, IF-OP50 + Glucose + IF, IH-OP50 + Glucose+IH, PA-OP50 + Glucose + PA. The data are shown as means SEM, with p < 0.05 indicating statistical significance. https://doi.org/10.6084/m9.Figureshare.24781494.
The effects of FBR extract on fat accumulation on C. elegans TG and FFA levels
Nile red fluorescence, TG, and FFA assays were used to determine the nematode fat deposition. Nile red fluorescent probe for intracellular lipids and proteins with hydrophobic domains (excitation/emission maxima ∼552/636 nm). Figure 4a shows that the worms in the treatment containing more fat ate more glucose than those in the NC treatment, implying that the worms’ diet was a major contributor to their increased fat content. Compared to the FBR-1741 and FA groups, less fat was found (Fig. 4a, b). Figure 4c demonstrates that FBR and FA inhibited TG in a dose-dependent manner, and the inhibitory effect was comparable to the Nile red and ORO staining findings. The genetic basis of fat metabolism has been extensively investigated using the C. elegans in vivo model. The metabolism, nutrient absorption, and reduction and deposition of fat were all carried out by the intestinal cells of C. elegans. TG enhancement is usually assessed as an endpoint when assessing food intake and energy expenditure. When administered to obese worms, FBR-1741 and FA increased their levels of free fatty acids. Normal, obese, FBR-174, FA, and orlistat worm groups had 3.86, 4.15, 9.23, and 6.59 nmol/mg free fatty acids, respectively (Fig. 5a). TG content increased in the obese model, then C. elegans treated with 10% glucose and FBR-1741 and FA decreased strongly (Fig. 5b). There are numerous possible explanations for this observation. The primary sources of free fatty acids are the breakdown of fat stores and dietary sources. Because of this, it is possible that an enhanced flux of free fatty acids in the worms is attributable to an abundance of these metabolites in the FBRs. Plant extract may also stimulate adipose enzymatic metabolism of endogenous fat, increasing plasma-free fatty acids. Compounds in plant extracts, in particular hormone-sensitive lipase, may catalyze the breakdown of adipose tissue triglycerides into fatty acids and glycerol. According to Rodrigues et al.39, specific phenolic compounds can promote lipolysis or the breakdown of triglycerides into glycerol and free fatty acids while inhibiting adipose growth and triglyceride production at the cellular level. Following Li et al.37, FA was analyzed with Oil Red O to investigate the significance of FA in suppressing fat accumulation. Furthermore, phenolic compounds can impact signaling pathways involved in adipogenesis. FBR-1741 can suppress lipid accumulation in C. elegans and extend its lifespan, according to moderate-to-strong connections between target lipid metabolism genes.

a Fatty acid levels. b Triglyceride level. The data are shown as means SEM, with p < 0.05 indicating statistical significance. https://doi.org/10.6084/m9.Figureshare.24781503.
Impact of FBR extracts on gene expression levels of C. elegans
There are many intricately controlled pathways involved in lipid metabolism in C. elegans. This study explored the levels of sterol regulatory element binding protein (sbp-1) and other important transcription factors that stimulate lipid production. In addition, we examined nuclear hormone receptor 49 (nhr-49), a transcription factor that promotes both fatty acid desaturation and β-oxidation, as a functional homolog of human peroxisome proliferator-activated receptors (PPARs)40. De novo lipogenesis-involved fatty acid desaturases (fat-4, fat-5, fat-6, and fat-7) are also positively regulated by nhr-4941. In addition, in C. elegans, Daf-16 is a key gene transcription factor that controls the insulin/IGF-1 regulatory system and promotes survival. Daf-16 is a gene primarily responsible for lifespan expansion, while it might also have an impact on fat accumulation in C. elegans18. Furthermore, hosl-1 specifies the C. elegans homolog of hormone-sensitive lipase42, which triggers the primary rate-limiting process in triglyceride breakdown (Supplementary Fig. 4).
Using qPCR, the FBR-1741 and metabolite samples were examined to determine how the fat-controlling genes in C. elegans had changed. As depicted in Figs. 6and 7, with the consumption of FBR-1741, the downregulation of sbp-1, fat-4, fat-5, fat-6, and fat-7 genes and upregulation of hosl-1 and daf-16 by N2 model-related lipid accumulation, lipolysis, and lifespan were observed (Fig. 6a–c). In our study, Daf-16 activity in neurons enhanced life expectancy by above 20%. In this study, we discovered that C. elegans, like mammals, accumulate fat when they consume excess calories from sugar. The mechanism for fat accumulation in C. elegans is nearly identical to that of mammals, suggesting that this nematode could be a time-saving and cost-effective alternative for investigating fat metabolism.

a Expression levels of genes involved in fat synthesis (FAT-4, FAT-6, FAT-7, and SBP-1). b Expression levels of genes involved in lipolysis (HOSL1). c Expression levels of genes involved in lifespan (Daf-16). d Heat map for the fatty acid metabolite profile. e Fat synthesis is responsible for fatty acid metabolites. The data are shown as means SEM, with p < 0.05 indicating statistical significance. https://doi.org/10.6084/m9.Figureshare.24781509.

This study provides an in-depth analysis of untargeted metabolomics and network pharmacology, highlighting the potential of fermented brown rice in enhancing anti-obesity efficacy. Through comprehensive research, we explore the complex interactions and biochemical pathways influenced by fermented brown rice, offering new insights into its therapeutic potential.
Effect of FBR extracts on fatty acid profiles of C. elegans
Human health is significantly affected by fatty acids, which consist of both saturated and unsaturated fatty acids. Saturated fatty acids are found primarily in palmitic, stearic, and arachidic acids; an excess of such acids can lead to atherosclerotic and cardiovascular disease43. The synthesis of unsaturated fatty acids depends on the genes that encode stearoyl-CoA desaturase (SCDs), such as fat-4, fat-5, fat-6, and fat-7. According to research, preventing the expression of fat-6 and fat-7, which encode the stearoyl CoA desaturating enzyme responsible for desaturating octadecyl saturated fatty acid, may minimize nematode fat accumulation44. Changes in metabolite levels occur in nearly all metabolic diseases. Several digestive enzymes that convert acetyl CoA to malonyl CoA and C16 palmitate regulate fat synthesis in C. elegans. This fluctuation may be restored to normal with the adoption of a nutritious daily diet (Supplementary Fig. 5a–c). Thus, this approach was used to show how metabolic alterations caused by obesity can be reversed by eating FBR-1741. Figure 6d shows that obese-induced FBR-1741 and FA treatment dramatically increased saturated fatty acid components, including palmitic acid (C16:0), stearic acid (C18:0), oleic acid (18:1), arachidic acid (20:4), eicosadienoic acid (20:5), and α-linoleic acid (PUFA) (Fig. 6d, Supplementary Fig. 5d, e). In addition, Qi et al.45 demonstrated that the ω-3 fatty acid α-linolenic acid (ALA) is able to increase the lifespan of C. elegans. Other treatments, such as positive control and metabolite diet groups, resulted in a significant decrease in saturated fatty acids. Serine, arginine, histidine, threonine, methionine, and glycolic acids also increased in FBR-1741- and FA-treated groups. According to Edwards et al.46, the levels of amino acids such as leucine, histidine, methionine, and tryptophan could significantly influence the lifespan of C. elegans. This finding provided further confirmation of our prior results, showing an increase in free fatty acid in worms after supplementation with FBR-1741, including metabolites for C. elegans. Results from both gene expression analysis and metabolomics assessment validate our prior finding that administering FBR-1741 and an FA-supplemented diet denoted that worm dramatically increased their lifespan (Supplementary Fig. 5f) and decreased lipid accumulation.
In this research, nine different types of brown rice were fermented with P. acidilactici MNL5 KCTC15156BP. Among the nine varieties, FBR-1741 had the strongest lipase inhibition, antioxidant activity, and phenolic phytochemical identification. In our previous investigation, we evaluated the biological activities of raw brown rice. In this study, we integrated omics data following systems biology principles to discover novel obesity therapeutic targets. However, FBR-1741 refers to specified phenolic compounds with potent lipase inhibition and antioxidant activity. Based on pathway enrichment studies, enhanced polyphenols may interfere with many pathways, including those implicated in obesity and obesity-related disorders, which may explain the possible anti-obesity impacts of FBR1741. The network-based pharmacological investigation of FBR-1741 found 17 compounds and 132 obesity-related target genes. Our results in the C. elegans N2 obese-induced paradigm reveal that FBR-1741 with ferulic acid has a higher survival rate and is a valuable indicator for assessing lipid reduction beyond dietary efforts. Ferulic acid was increased by FBR-1741, which may have a causal relationship with increased longevity and decreased lipid levels. In addition, FBR-1741 prolonged C. elegans lifespan and reduced fat accumulation, validating gene expression for fatty acid metabolites. However, additional research is needed to explain the fermented material for the in vivo C. elegans metabolomic study. This research provides a better comprehension of the synergistic effects of microorganisms. In addition, the findings of this study will be useful for future research using obesity models in mice to investigate the effect of FBR-1741 on gut microbiota and obesity correlation studies. In addition, we can use these considerations to develop a fermented brown rice product to combat obesity.
Source link

